Характеристика археоптерикса таблица позвоночник

Археоптерикс – вымершее животное размером с ворону, систематическое положение которого до конца не определено. В зависимости от научного взгляда, одни палеонтологи его относят к динозаврам, другие к птицам (одно из названий данных археоптериксу – птица пресмыкающееся).
История находки
- За всю историю палеонтологии удалось обнаружить и описать 12 экземпляров археоптерикса.
- Впервые нового представителя древнего мира описал в 1861 году Герман фон Майер, на основе отпечатка единственного пера в породах района Зольнхофена нынешней Германии. В том же году в тонкослоистых известняках вблизи Лангенальтайма нашли неполный скелет существа, описанный в 1863 году Р. Оуэном под названием «Лондонский экземпляр». Реконструированный образец стал прообразом рода Archaeopteryx (лат.).
До конца XX века ученые описали еще 8 окаменелостей археоптерикса, найденных в Зольнхофенских горных породах:
- Берлинский. Останки найдены в 1876 или 1877 году Й. Дёрром. Я. Нимейер выменял их у него за корову. Восстановленный образец классифицирован в 1884 как новый вид Archaeopteryx siemensii. (Встречаемое в литературе название этого экземпляра – Archaeornis)
- Харлемский. Найден в 1855. В 1875 Мейер по ошибке принял его за вид птеродактиля. В 1970 Д. Остром детально изучил образец и причислил его к роду археоптериксовых.
- Айхштетский. Этот экземпляр был найден в районе деревни Воркерсцелль в 1951 году, но описан только в 1974 из-за этической дилеммы по поводу ценности находки между владельцем Ф. Фрейем и ученым К. Мейером.

- Максбергский. Скелет без головы и хвоста нашли в 1956, через три года его описал профессор Ф. Хеллер.
- Зольнхофенский. Добыт в 1970 году и по ошибке отправлен в коллекцию Ф. Мюллера под видом останков компсогната. Позднее при открытии нового музея, проводившие оценку образца П. Веллнхофер и Г. Фиоль, установили принадлежность экспоната к археоптериксу.
- Дайтингский археоптерикс отличался от всех образцов местом находки. Окаменелости нашли в 1990 году в отложениях Дайтинга, которые на десятки тысяч лет моложе других находок. Спустя 20 лет оригинал описал Р. Альберсдорфер, в честь которого в 2018 году вид классифицировали как A.
- Мюнхенский. При первичном описании в 1993 образец стал единственной окаменелостью с хорошо сохранившейся грудиной. Впоследствии П. Веллнхофер опроверг поспешный вывод, так как при исследовании в ультрафиолетовом диапазоне грудина оказалась сместившимся участком левой воронковидной кости.
- Фрагментарный экземпляр обнаружил в 1997 году доктор М. Маузер в коллекции частного владельца, имя которого не разглашается. Официально образец не классифицирован, поэтому к роду археоптериксов его зачислили предварительно.
В начале XXI века таксон пополнился двумя новыми экземплярами:
- Термополиский. Плиту с прекрасно сохранившимся отпечатком скелета с местами прикрепления перьев к телу нашли в частной экспозиции в 2005.
- Двенадцатый образец в 2010 году нашел немецкий коллекционер, но мир палеонтологии узнал о находке лишь в 2014. Это старейший из археоптериксов, так как слои породы с окаменелостями археоптерикса на 200-300 тыс лет старше Зольнхофенского известняка.
Виды археоптерикса
На протяжении всего периода находок окаменелых костей археоптерикса первоначально практически каждый из них относили к отдельному роду или виду.
Разногласия возникали из-за существенных отличий в анатомии образцов. В дальнейшем все особенности стали связывать с половым диморфизмом и разными возрастными стадиями древнего существа.
Официально признается три вида:
- siemensii (альтернативное название A. bavarica);
- lithographica (синонимы A. macrura, A. oweni);
- albersdoerferi.

Строение скелета
В большинстве работ палеонтологов археоптерикс описывается как первая птица произошедшая от ящеротазовых динозавров. Некоторые исследователи считают его потомков тероподов. Но на самом деле точное систематическое положение существа до конца не определено, есть версия, что археоптерикс – переходная форма между рептилиями и птицами. Причиной двойственной принадлежности вида к древним пресмыкающимся и птицам стало наличие анатомических особенностей, характерных для каждой из групп.
Строение позвоночника и верхних конечностей
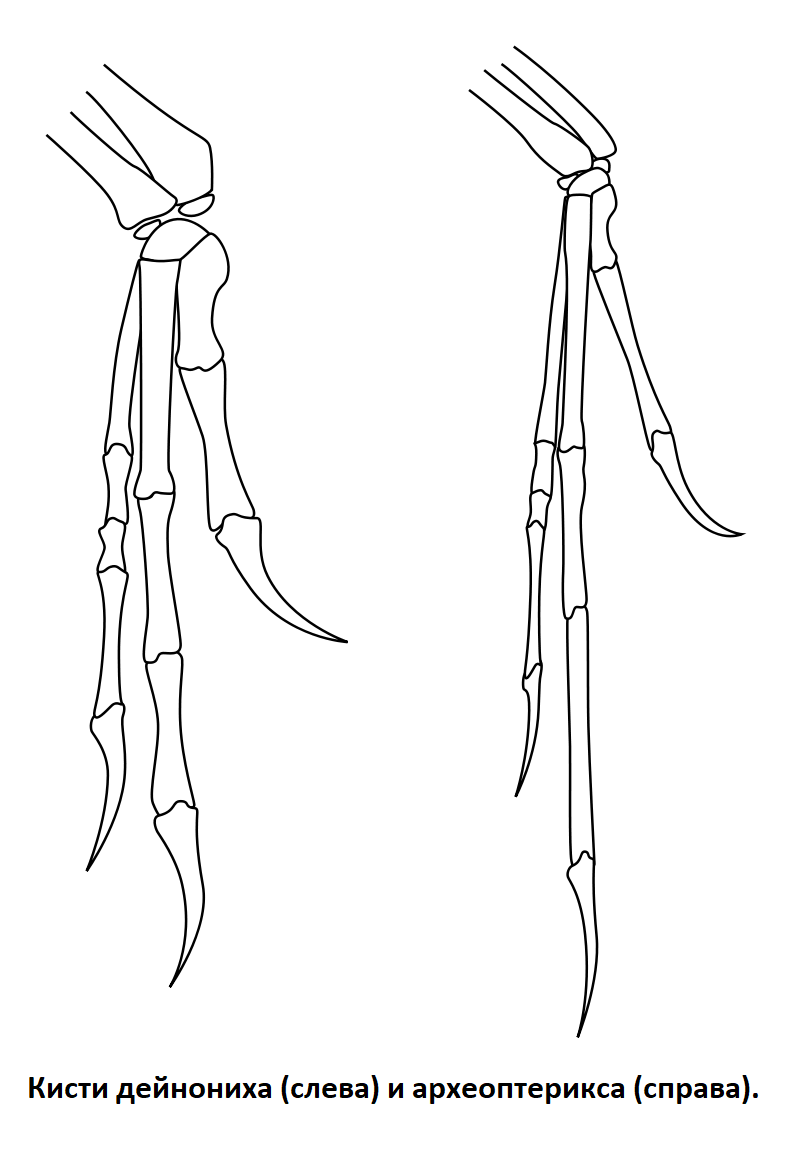 У археоптерикса передние конечности были длинными, с тремя хорошо развитыми пальцами. Формула фаланг: 2-3-4. Заканчивались пальцы длинными загнутыми когтями. Кости пясти и запястья не срослись, полулунная кость составляла часть запястья.
У археоптерикса передние конечности были длинными, с тремя хорошо развитыми пальцами. Формула фаланг: 2-3-4. Заканчивались пальцы длинными загнутыми когтями. Кости пясти и запястья не срослись, полулунная кость составляла часть запястья.
Шейные позвонки археоптерикса двояковогнуты: вогнуты сзади и спереди, что присуще динозаврам семейства дейнонихозавров.
В крестовом отделе позвоночника у археоптерикса пять несросшихся позвонков (у птиц часть поясничных, крестцовые и начальные хвостовые позвонки срастаются в одну кость – сложный крестец, срастаются тазовые кости, а оставшиеся хвостовые позвонки срастаются, образую кость – пигостиль). Подвздошная, седалищная и лобковая кости археоптерикса не образуют единую кость, дистальные концы лобковых костей соединяются и образуют лобковый симфиз, а хвост у археоптерикса длинный и состоит из 21-23 отдельных позвонков.
У археоптерикса были найдены вентральные ребра (гастралии), что характерно для пресмыкающихся, без крючковидных отростков (эти отростки у птиц обеспечивали прочность грудной клетки). Ключицы археоптерикса срастались и образовывали вилочку, что характерно как для птиц, так и для динозавров.
Строение нижних конечностей
Голень состоит из большой и малой берцовых костей, примерно одинаковой длины. Часть костей предплюсны и плюсны не срослись, а это значит, что у археоптерикса отсутствовала цевка.
Большой палец противопоставлен остальным, что является характерным для птиц . На втором, самом длинном пальце задней конечности, рос длинный коготь, что присуще динозавром семейства дейнонихозавров.
Строение черепа
На предчелюстных, челюстных и зубных костях археоптерикса росли мелкие конические зубы, как у современных крокодилов. Предчелюстные кости не срастались. У нижней и верхней челюсти отсутствовал роговой чехол, а значит клювом археоптерикс не обладал. На затылке, сзади, у археоптерикса расположено отверстие, соединяющее позвоночный канал и череп.
Мозг археоптерикса такого же объема, что у современных птиц и в три раза больше, чем у рептилий того же размера. Полушария мозга археоптерикса не окружают обонятельные трактаты и они пропорционально меньше, чем у современных птиц.
Мозжечок динозавра больше, чем у тероподов, но меньше, чем у современных птиц. Зрительные доли мозга расположены спереди. Полет археоптерикса был невозможен без увеличенных зрительных долей и мозжечка.
Положение переднего и бокового полукружных каналов у археоптерикса типично для архозавров, но передний канал длиннее и изогнут в обратном направлении.
Перья археоптерикса

Именно это окаменевшее перо археоптерикса с которого началась история пернатых динозавров
Лондонский и берлинский экземпляры археоптерикса сохранили отпечатки перьевого покрова. Оперение – это ключевая особенность, по которой археоптерикса относят к птицам.
Перья археоптерикса, как и у современных птиц, подразделялись на маховые, рулевые и контурные. Маховые и рулевые перья археоптерикса имеют стержень пера, бородки 1-го порядка и бородки 2-го порядка. Для маховых перьев характерно ассиметричное строение опахал, что типично для современных птиц, способных к полёту. Отличие перьевого покрова современных птиц от археоптерикса – наличие крылышка (подвижного пучка перьев на большом пальце передних конечностей).
Способность к полету
Строение перьевого покрова (ассиметричное строение маховых перьев) подтверждает тот факт, что археоптерикс был способен летать. Но ученые еще не пришли к общему мнению о том был ли это полет примитивным (планирующим), или более сложным (машущим), как у поздних птиц.
При оценке нагрузки на крыло ученые утверждают, что этот древний ящер был способен только к планирующему полету, так как не мог поднять крыло выше уровня спины, что является необходимым условием для замаха.
Другие исследования указывают что форма тела и размер крыла были нетипичны для планирующих птиц, а вилочка и коракоид служили для прикрепления мышц, двигающих крыло.
Интересным представляется гипотеза, что археоптерикс мог совершать небольшие движения крыльями без замаха. Эти движения совершались во время планирующего полета.
Образ жизни
Одни учёные склоняются к тому, что археоптерикс вёл наземный образ жизни, другие считают, что древесный. Скорее всего, он искал пищу и в кустарниках, и на земле. Мелкую добычу он хватал челюстями, для крупной пускал в ход когти. Судя по строению глазниц динозавр вел ночной образ жизни.
Признаки принадлежности археоптерикса к динозаврам:
-
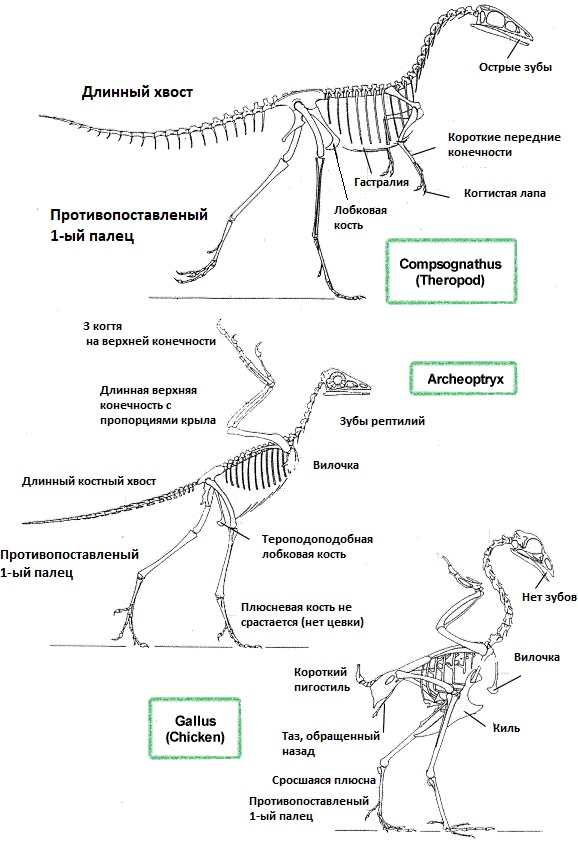 отсутствие птичьего клюва и цевки (кость между голенью и пальцами у птиц);
отсутствие птичьего клюва и цевки (кость между голенью и пальцами у птиц); - остроконечные зубы в предчелюстных, челюстных и зубных костях;
- расположение большого затылочного отверстия сзади черепа;
- вентральные ребра без крючковидных отростков;
- полноценно развитые пальцы на конце длинных передних конечностей (фаланговая формула 2-3-4 соответствует дромеозаврам);
- отсутствие срастания подвздошных костей и крестца в единую кость;
- наличие в крестцовом отделе 5 позвонков, как у дейнонихозавров;
- сильно изогнутые когти на пальцах.
Признаки птиц у археоптерикса:
- оперение – ключевая особенность, по которой археоптерикс не считается динозавром;
- увеличенный объем мозга по сравнению с рептилиями того периода;
- срастание ключиц в вилочку.
Музеи, где представлены скелеты археоптерикса
Большинство скелетов археоптериксов экспонируется на территории Германии в:
- Музее естествознания в Берлине;
- Музее динозавров в природном парке Альтмюльталь в Денкендорфе;
- Музее бургомистра Мюллера в Зольнхофене;
- Палеонтологическом музее Jura-Museum в Айхштете;
- Палеонтологическом музее Мюнхена.
Остальные реконструированные экспонаты выставлены в:
- Центре динозавров в Термополисе (штат Вайоминг, США)
- Музее Тейлора в Харлеме (Нидерланды);
- Лондонском Музее естествознания (Англия).
Для палеонтологической истории XIX века археоптерикс стал подтверждением теории эволюционного развития птиц от динозавров, описанной в книге Ч. Дарвина «Происхождение видов». Со временем это предположение поставили под сомнение. В юрских отложениях были найдены более убедительные образцы древних существ, доказывающих происхождение птиц от пресмыкающихся.
Предыдущая
ХищникиКетцалькоатль | Птерозавр Европы
Следующая
ХищникиГерреразавр (эрреразавр) | Динозавр Южной Америки (Аргентина)
Источник
Тип урока: изучение нового материала.
Форма урока: фронтальная, индивидуальная.
Методы обучения: эвристический метод, объяснительно-иллюстративный, практический, наглядный.
Оборудование: Презентация “Доказательства эволюции””, компьютер, мультимедийный проектор.
Задачи урока:
- раскрыть научное значение обнаружения в развитии зародышей следов далекой истории систематических групп;
- дать оценку биогенетического закона Ф. Мюллера и Э. Геккеля как эмбриологического доказательства;
- выяснить значение для науки ископаемых переходных форм как палеонтологических доказательств, изучить сравнительно-анатомические (морфологические), биогеографические доказательства эволюции.
Ход урока
Презентация.
I. Изучение нового материала (тема урока на слайде 1).
1. Палеонтологические доказательства (слайд 2).
Палеонтология (от греч. палео – древний; онтос – существо; логос – учение) – наука о животных и растениях прошлых геологических эпох, изучаемых по ископаемым остаткам и следам жизнедеятельности. Основатели палеонтологии: Ж. Кювье, Ж.-Б. Ламарк, А. Броньяр. Термин “палеонтология” был предложен в 1822 г. А. Бленвилем. Основы современной эволюционной палеонтологии заложил В. О. Ковалевский.
Палеонтология решает следующие задачи:
- выявление ранних этапов эволюции жизни;
- выявление обособления стволов органического мира;
- выявление основных этапов развития органического мира;
- выявление событий на рубежах основных подразделений истории Земли.
Методы палеонтологии:
- палеонтологический, заключающийся в расчленении осадочных толщ, определении их относительного возраста и установлении сходства между слоями разных регионов по сохранившимся в них ископаемым остаткам;
- радиоактивный, основанный на изучении природного распада радиоактивных элементов, содержащихся в горных породах; позволяет вычислить абсолютный возраст горных пород и останков ископаемых организмов.
Палеонтология предоставляет следующие данные в пользу эволюции:
Сведения об ископаемых переходных формах (определение переходных форм было дано выше), которые не дожили до наших дней и присутствуют только в виде ископаемых останков. Примерами ископаемых переходных форм являются: древние кистеперые рыбы, семенные папоротники, псилофиты, зверозубый ящер, археоптерикс и др. Существование переходных форм между различными типами и классами показывает, что постепенный характер исторического развития свойственен не только низшим систематическим категориям (видам, родам, семействам), но и высшим категориям и что они также являются закономерным результатом эволюционного развития (Слайды 3, 4).
Сведения о филогенетических (эволюционных) рядах (Слайд 5), которые не только являются прекрасной иллюстрацией эволюции, но и позволяют узнать причину эволюции отдельных групп организмов. В этом плане интересны исследования В. О. Ковалевского истории развития лошадей. Современные однопалые животные произошли 60-70 млн. лет назад от мелких пятипалых всеядных предков размером с собаку. В ходе приспособлений к жизни в степной равнинной местности с твердым грунтом пятипалая конечность превратилась в однопалую, утратив боковые пальцы. Этот пример предельною развития признака В. О. Ковалевский назвал “принципом кульминирования”. Одновременно с изменением строения конечностей у предков современных лошадей увеличивались размеры тела, изменялась форма черепа, усложнялось строение зубов, шло возникновение пищеварительного тракта, свойственного травоядным. Филогенетические ряды убедительно показывают, что эволюция в целом носит приспособительный характер.
Таким образом, несмотря на неполноту, палеонтологическая летопись предоставляет убедительные доказательства эволюции.
2. Сравнительно-анатомические (морфологические) доказательства (слайд 6).
Сравнительная морфология – биологическая дисциплина, изучающая закономерности строения и развития органов и их систем путем сопоставления организмов разных систематических групп. У истоков сравнительной морфологии как науки стояли Ж. Кювье, Ж. Сент-Иллер и другие ученые.
К доказательствам эволюции этой группы относятся:
1) наличие в современной флоре и фауне переходных форм, являющихся родоначальниками нескольких систематических групп и сочетающих в себе признаки этих групп организмов (зеленая эвглена, сидячая асцидия, латимерия, утконос, ехидна, ланцетник и др.). Переходные формы свидетельствуют о преемственности в эволюции и о том, что низшие дали начало высшим;
2) наличие в пределах класса, типа гомологичных органов, образований, сходных друг с другом по общему плану строения, положению в теле и возникновению в процессе онтогенеза (Слайды 7-10). Гомология связана с наличием у разных видов одинаково действующих наследственных факторов (так называемых гомологичных генов), доставшихся от общего предка. Например, скелет конечностей всех четвероногих выводится из основной пятипалой схемы, хотя сами конечности могут выполнять различные функции. Гомологии противостоит аналогия. Аналогичные органы – это образования с одинаковыми функциями, но с различным планом строения, онтогенезом и происхождением. Например, копательные конечности у крота и медведки, почки млекопитающих и мальпигиевы сосуды насекомых и др.;
3) наличие рудиментов (от лат. rudimentum – зачаток, первооснова) – сравнительно упрощенных, недоразвитых, по сравнению с гомологичными структурами предков, органов, утративших свое основное значение в организме в ходе эволюционного развития (Слайд 11-13). Рудименты закладываются во время зародышевого развития организма, но полностью не развиваются. Они встречаются у всех особей данного вида. Например, малая берцовая кость у птиц, тазовый пояс у кита, глаза у роющих животных и др.;
4) наличие атавизмов (от лат. atavus – предок), признаков, появляющихся у отдельных особей данного вида, которые существовали у отдаленных предков, но были утрачены в процессе эволюции. Например, изредка появляющиеся у китов задние конечности, хвостатость у человека и др. Атавизмы являются проявлением эволюционной памяти о предках. Причины их появления заключаются в том, что гены, ответственные за данный признак, сохраняются в эволюции данного вида, но их действие при нормальном развитии блокируется генами-репрессорами. Через много поколений в онтогенезе отдельных особей по отдельным причинам блокировка снимается и признак проявляется вновь.
Таким образом, данные сравнительной морфологии убедительно свидетельствуют об изменяемости органического мира.
Примеры:
а) формы, сочетающие в себе признаки нескольких крупных систематических единиц (эвглена зеленая);
б) связь между разными классами животных свидетельствует об общности их происхождения. Яйцекладущие (утконос, ехидна) по ряду особенностей своей организации промежуточны между рептилиями и млекопитающими;
в) строение передних конечностей некоторых позвоночных (Слайд 14) (ласт кита, лапа крота, крыло птицы, рука человека, лапа крокодила имеют кости плеча, предплечья, кисти, только они разной формы и размеров).
3. Эмбриологические доказательства эволюции.
А) Предмет эмбриологии.
Эмбриология (от греч. эмбрион – зародыш и логос – учение) – в узком смысле наука о зародышевом развитии, в широком смысле – наука об индивидуальном, развитии организмов (об онтогенезе). Эмбриология животных и человека изучает предзародышевое развитие (оогенез и сперматогенез), оплодотворение, зародышевое развитие, личиночный и постэмбриональный периоды индивидуального развития.
Эмбриология в зависимости от задач делится на: общую, сравнительную, экспериментальную, популяционную и экологическую.
Б) К данным эмбриологии, являющимися доказательствами эволюции, относят (слайд 15):
Закон зародышевого сходства Карла Бэра, который гласит: “Эмбрионы обнаруживают, уже начиная с самых ранних стадий, известное общее сходство в пределах типа” (работа учащихся с рисунками, иллюстрирующими ход зародышевого развития представителей различных классов типа Хордовые (Слайд 16)). У всех хордовых на ранних стадиях развития закладывается хорда, возникает нервная трубка, в переднем отделе глотки образуются жабры и т. д. Сходство зародышей свидетельствует об общности происхождения данных организмов. По мере развития зародышей черты их различия выступают все более явственно. К. Бэр первым обнаружил, что в ходе эмбрионального развития сначала появляются общие признаки типа, затем последовательно класса, отряда и, наконец, вида.
Расхождение признаков зародышей в процессе развития называют эмбриональной дивергенцией, и она объясняется историей данного вида.
Биогенетический закон Геккеля-Мюллера, указывающий на связь индивидуального (онтогенеза) и исторического (филогенеза) развития. Этот закон был сформулирован в 1864-1866 гг. немецкими учеными Ф. Мюллером и Э. Геккелем. В своем развитии многоклеточные организмы проходят одноклеточную стадию (стадию зиготы), что может рассматриваться как повторение филогенетической стадии первобытной амебы. У всех позвоночных закладывается хорда, которая далее замещается позвоночником, а у их предков хорда оставалась всю жизнь. В ходе эмбрионального развития птиц и млекопитающих появляются жаберные щели в глотке. Этот факт можно объяснить происхождением этих наземных животных от рыбообразных предков. Эти и другие факты и привели Геккеля и Мюллера к формулировке биогенетического закона. Он гласит: “Онтогенез есть краткое и быстрое повторение филогенеза, каждый организм в индивидуальном развитии повторяет стадии развития предков”. Образно говоря, всякое животное во время своего развития взбирается по собственному родословному древу. Однако онтогенез не так уж точно повторяет филогенез. Поэтому повторение стадий исторического развития вида в зародышевом развитии происходит в сжатой форме, с выпадением ряда этапов. Кроме того, эмбрионы имеют сходство не со взрослыми формами предков, а с их зародышами.
4. Биогеографические доказательства.
Биогеография (от греч. био – жизнь, гео – земля, графа – пишу) – наука о закономерностях распространения по земному шару сообществ живых организмов и их компонентов – видов, родов и других таксонов. В состав биогеографии входят зоогеография и ботаническая география. Основные разделы биогеографии стали оформляться в конце XVIII и в 1-й половине XIX веков, благодаря многочисленным экспедициям. У истоков биогеографии стояли А. Гумбольдт, А. Р. Уоллес, Ф. Склетер, П. С. Паллас, И. Г. Борщов и др. Современная биогеография включает в себя следующие разделы: ареалогическая, региональная, экологическая и историческая биогеография.
К данным биогеографии, являющимися доказательствами эволюции (слайд 17), относятся следующие:
Особенности распространения животных и растений по разным континентам, как яркое свидетельство эволюционного процесса. А. Р. Уоллес, один из выдающихся предшественников Ч. Дарвина, привел все сведения о распространении животных и растений в систему и выделил шесть зоогеографических областей (работа учащихся с картой зоогеографи-ческих областей мира): 1) палеоарктическую (Европа, Северная Африка, Северная и Средняя Азия, Япония); 2) неоарктическую (Северная Америка); 3) эфиопскую (Африка к югу от Сахары); 4) индомалайскую (Южная Азия, Малайский архипелаг); 5) неотропическую (Южная и Центральная Америка); 6) австралийскую (Австралия, Новая Гвинея, Новая Зеландия, Новая Каледония). Степень сходства и различия флор и фаун между разными биогеографическими областями неодинакова. Так, палеоарктическая и неоарктическая области, несмотря на отсутствие между ними сухопутной связи, обнаруживают значительное сходство флор и фаун. Животный и растительный мир неоарктической и неотропической областей, хотя между ними и имеется сухопутный Панамский перешеек, сильно отличаются друг от друга. Чем это можно объяснить? Это можно объяснить тем, что некогда Евразия и Северная Америка входили в состав единого континента Лавразии и их органический мир развивался совместно. Сухопутная связь между Северной и Южной Америкой, напротив, возникла относительно недавно, и их флоры и фауны долгое время развивались обособленно. Особняком стоит органический мир Австралии, которая обособилась от Южной Азии более 100 млн. лет назад, и лишь в ледниковый период сюда через Зондский архипелаг перебрались немногие плацентарные – мыши и собаки. Таким образом, чем теснее связь континентов, тем более родственные формы там обитают, чем древнее изоляция частей света друг от друга, тем больше различия между их населением.
Особенности фауны и флоры островов также свидетельствуют в пользу эволюции. Органический мир материковых островов близок к материковому, если отделение острова произошло недавно (Сахалин, Британия). Чем древнее остров и чем значительнее водная преграда, тем больше отличий в органическом мире этого острова и близлежащего материка (Мадагаскар). Органический мир вулканических и коралловых островов беден и является результатом случайного занесения некоторых видов, способных перемещаться по воздуху.
II. Закрепление знаний.
Лабораторная работа “Изучение палеонтологических доказательств эволюции”.
Заполнение учащимися таблиц по вариантам (можно пользоваться учебниками зоологии и общей биологии).
Характеристика археоптерикса
| Части тела и жизненные процессы | Признаки пресмыкающихся | Признаки птиц |
| Позвоночник | Позвонки несросшиеся, спинные ребра прямые, без отростков; брюшные ребра | Ключицы срослись в вилочку |
| Грудная клетка | Грудная кость без киля | |
| Череп | Небольшой по объему; челюсти с зубами, нет рогового чехла клюва | |
| Передние конечности | Кости в основном без полостей, кисть вооружена когтями | Крылья |
| Хвост | Длинный, состоящий из 20 отдельных позвонков | |
| Задние конечности | Наличие цевки | |
| Кожный покров | Чешуи | Наличие перьев |
| Общий вид | Величиной с голубя, напоминал птицу | |
| Образ жизни | Лазающий | Способность планировать |
Источник